Breathing, Aerobic Conditioning and Gas Consumption
Breathing, Aerobic Conditioning and Gas Consumption
BY JOHNNY E. BRIAN, JR., M.D.
Breathing is an essential part of life. For divers, breathing is of special interest, as it determines the duration of our gas supply. The majority of lung ventilation is regulated without conscious input, being governed by the need for oxygen and the elimination of carbon dioxide (CO2). However, with aerobic conditioning, divers can influence lung ventilation and reduce their gas consumption. Diving is a sport focused on gas consumption. Understanding how aerobic conditioning affects the respiratory system allows us to become better, more informed divers. Aerobic conditioning is a complex, multifactoral process, of which the respiratory system is only a part. For the purposes of this article, I will address only the respiratory component of aerobic conditioning. To understand these complex events, the best place to start is with basic respiratory physiology.
RESPIRATORY PHYSIOLOGY At rest and during exercise, we breathe to deliver oxygen to and eliminate CO2 from our bodies. Tidal volume (TV) is the amount of gas inhaled and exhaled with each breath, and averages 6 to 8 milliliters (ml) per kilogram (kg) of body weight (see Table 1). In a typical 70 kg diver, this results in a TV of approximately 500 ml. (Divers more familiar with English units should consult the appendix to convert English to metric units.) At rest, we breathe 12 to 16 times a minute, which results in a minute ventilation (MV) of 6 to 7 liters (L) per minute (TV x rate). At rest, when oxygen consumption is minimal, the need to eliminate carbon dioxide (CO2) controls ventilation. When CO2 dissolves in water, it forms carbonic acid (H2O + CO2 = H2CO2), which reduces the pH (a measure of acidity and alkalinity) of the solution. Since our bodies are principally composed of water, changes in CO2 profoundly influence the pH of our bodies. Under resting conditions, what drives our ventilation is CO2-mediated change in brain pH. Normal arterial partial pressure of CO2 is 35 to 45 mmHg. The respiratory control system in the brain is very sensitive to CO2-mediated change in brain pH. In healthy individuals at rest, arterial partial pressure of oxygen averages 95 mmHg. To produce significant respiratory drive, arterial partial pressure of oxygen must be less than 60 mmHg, rare in healthy individuals. Although the arterial CO2 directly controls ventilation, this does not imply that ventilation is independent from oxygen consumption. On an average, 80% of oxygen consumed is converted to CO2 (the remaining 20% is converted to water), so that as oxygen consumption increases, CO2 production also increases, which drives ventilation to supply more oxygen and eliminate more CO2. At rest, our body's metabolism is minimal, and the cells of our body carry on basic metabolic processes to sustain themselves. The actual type of cell is less important than the number of cells. Most studies of aerobically fit individuals indicate that, at rest, oxygen consumption is not different than in unconditioned individuals when body mass is considered. Not surprisingly, minute ventilation at rest also is not different between conditioned an unconditioned individuals. Thus, at rest, the total amount of gas we breathe is primarily determined by body mass. At rest, any given gas supply will last longer for a smaller individual.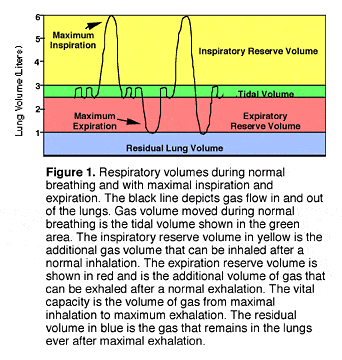 |
| Figure 1 |
Ventilation increases with exercise, but factors responsible for regulating ventilation during exercise are poorly understood. With the onset of exercise, ventilation increases before arterial CO2 can increase; in general, arterial O2 does not decrease with exercise. The overall regulation of ventilation during exercise likely results from integration of increased input from peripheral receptors (movement of joints, muscles), and central input (we know we are exercising, and must increase our ventilation). During light and moderate levels of exercise, arterial partial pressures of O2 and CO2 are maintained at resting values. During exercise approaching maximal values, arterial CO2 decreases. During extreme exercise in elite athletes, arterial O2 partial pressure may also decrease.
In an aerobically conditioned individual, light and moderate levels of exercise result in an increase in tidal volume and little to no increase in the respiratory rate. We increase our tidal volume by using some of the lung reserve volume, principally the inspiratory reserve volume. Taking deeper breaths is advantageous, as deeper breaths increase the efficiency of respiration. With each breath, some gas always remains in the airways that conduct gas to the gas exchange portion of the lungs. The gas in the conducting airways does not undergo gas exchange, and thus is "wasted" or "dead space" ventilation. In essence, the oxygen in dead space gas is unavailable for gas exchange, and is thus "wasted" ventilation. On an average, dead space ventilation averages 2.2 ml/kg, which would be 150 ml in our example 70 kg person. This means at rest, 150 ml of each 500 ml breath, or 30% of inhaled gas, does not undergo gas exchange. For a 7 L minute ventilation, this means that 2.1 L of gas breathed each minute does not contribute to gas exchange.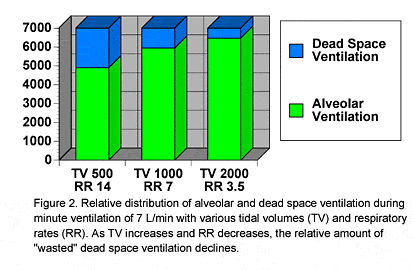 |
| Figure 2 |
Dead space ventilation is relatively fixed and does not change when tidal volume changes. Larger breaths, however, increase breathing efficiency by reducing the percentage of dead space ventilation. For a 1000 ml breath, the dead space ventilation would remain 150 ml, but the percent dead space ventilation would decrease to 15%. In this case, the amount of gas breathed per minute that does not undergo gas exchange is reduced to 1.05 L. In Figure 2, a minute ventilation of 7 L/min is divided into increasing tidal volumes with decreasing respiratory rates, so that the total ventilation per minute remains constant. With larger and larger tidal volumes, the percentage of dead space ventilation progressively decreases. The important effect of reducing dead space ventilation is to allow more inhaled gas to undergo exchange with the blood, and increase the oxygen available to blood. Taking fewer, larger breaths is more efficient than fewer, smaller breaths, as oxygen used by the respiratory muscles is only very slightly increased for larger breaths. The increase in breathing efficiency from larger and larger breaths is eventually offset by the increasing requirement for oxygen to support the work of breathing. At some point, the metabolic cost of taking a larger breath is not offset by increased oxygen delivery into the system.
Almost everyone will arrive at the most efficient combination of respiratory rate and tidal volume during the course of aerobic training. During maximal exercise, elite athletes usually have tidal volumes of approximately 60% of vital capacity. The important point for divers is that taking deeper breaths is key for increased ventilatory efficiency during exercise. Restriction of our ability to take deeper breaths, as from restrictive regulators, suits, or harnesses, will reduce our ability to maximize ventilatory efficiency.One of the important respiratory adaptations of aerobic cond-itioning is that less minute ventilation is required at any given exercise intensity (oxygen consumption). In other words, fit people will always breathe less gas to support a given level of exercise compared to unconditioned people. Resting oxygen consumption is approximately 3.5 ml/kg/min, or about 250 ml/min in a 70 kg individual. The ventilatory equivalent is the volume of gas breathed to extract a given amount of oxygen. At rest, and during submaximal exercise, the ventilatory equivalent averages 20 to 25 L of gas for each 1 L of oxygen consumed. One of the hallmarks of aerobic conditioning is that the ventilatory equivalent is reduced over the course of training. With progressive aerobic training, less gas is breathed to support a given level of exercise. This means that less respiratory work is required to maintain a given level of exercise, and less oxygen will be consumed by the respiratory muscles. With aerobic conditioning, there are other important cardiac and circulatory adaptations that also affect the efficiency of oxygen use. (The cardiovascular component of aerobic conditioning will be addressed in a separate article.)
Aerobic conditioning is obviously much more than learning to take deeper breaths. There are other adaptive changes in the respiratory system that influence gas utilization. We are dependent on respiratory muscles to move gas in and out of the respiratory system. Our ability to sustain exercise is determined in part by the ability of the respiratory muscles to sustain work and gas movement. Aerobic conditioning increases the aerobic capacity of respiratory muscles, just as it does for skeletal muscles. During aerobic conditioning, the oxidative enzyme capacity of respiratory muscles increases, which increases the ability of the muscles to metabolize oxygen. This means that the point at which respiratory muscles convert from aerobic to anaerobic metabolism is moved to a higher work level. Thus, aerobic conditioning of respiratory muscles allows greater gas movement per minute without accumulation of blood lactate derived from the respiratory muscles. Aerobic conditioning of respiratory muscles also increases the strength of the muscles, which allows generation of larger, more sustained pressure gradients to move gas in and out of the airway. These changes together allow more sustained aerobic activity of the respiratory muscles, which helps protect them against fatigue. Swimming offers a unique advantage in enhancing respiratory muscle function. Because the respiratory muscles must work against the added load of water compressing the chest, the respiratory muscles must generate more force with each breath. Thus, the respiratory muscles are strengthened to an added degree during swim training.
Lung volume is related to body size, with larger individuals having larger lungs. Lung volume also determines the surface area for gas exchange. One might think that having a larger lung volume and surface area for gas exchange would be advantageous during aerobic exercise. However, this does not appear to be the case, as marathon runners have lung volumes no different than size-matched sedentary individuals. The greater lung surface area found in larger individuals is likely necessary to support the needed gas exchange for a larger body mass. Swimmers, however, do have larger lung volumes than size-matched non-swimmers. Because of the restriction imposed on ventilation by swimming, it is advantageous to have a larger lung volume, which allows more oxygen to be held in the lungs for exchange during the periods between breaths. The larger lung volumes found in swimmers may be an adaptive change to breath-holding. Measurement of lung volumes in individuals who participate in breath-hold diving demonstrates larger lung volumes after one year of repetitive breath-hold diving. In compressed gas diving, if off-gassing during decompression is diffusion-limited, then a larger surface area for gas exchange might be advantageous during decompression. This has not been tested experimentally, and would likely be difficult to evaluate due to the multifactorial nature of decompression. However, most decompression models are perfusion limited, and not diffusion limited. The respiratory, cardiovascular and muscular systems function as an integrated system during exercise. It should not be surprising that the reduced respiratory requirements that occur with aerobic training appear to be specific to the type of training. This means that the reduced ventilatory requirements that occur with aerobic training by running, for example, do not fully transfer to other types of exercise. Thus, to obtain the maximal benefits from aerobic conditioning, one should include a variety of exercise types in their training program. To train for diving, swimming has obvious advantages, as it conditions the muscle groups used during diving, and has added benefits in conditioning of the respiratory muscles. SUMMARYThe respiratory effect of aerobic conditioning includes both a "learned" and a physiologic component. We innately learn to breath more efficiently during aerobic conditioning, and through continued aerobic exercise arrive at a breathing pattern that is most efficient. With conditioning, our respiratory muscles develop increased aerobic capacity, which increases the threshold for the onset of anaerobic metabolism and lactate accumulation in the muscles. There is no magic to aerobic conditioning, just the time and effort.
All rights reserved.
